Learning Objectives
6 objectivesBy the end of this note, you should be able to:
- Know the properties of gas exchange surfaces: surface area to volume ratio, thickness, and concentration difference.
- Understand how diffusion rate depends on these properties using Fick’s Law.
- Understand how mammalian lung structure is adapted for rapid gas exchange.
- Know the structure and properties of cell membranes.
- Understand how the fluid mosaic model interprets data on membrane structure.
- Investigate the effect of alcohol and temperature on membrane permeability.
Properties of Gas Exchange Surfaces
Efficient gas exchange surfaces share three key properties: a large surface area to volume ratio, a thin exchange surface, and a steep concentration difference between the two sides.
A large surface area provides more space for diffusion to occur simultaneously. Single-celled organisms such as Amoeba rely on direct diffusion across their cell surface because they have a high surface area to volume ratio. As body size increases, volume rises much faster than surface area, so larger organisms cannot rely on diffusion alone. They develop specialised exchange surfaces such as gills, alveoli, and root hairs to compensate.
A thin exchange surface shortens the diffusion pathway. Most are only one cell thick, like the alveolar wall in human lungs. This minimises the distance gases must travel from air to blood.
A steep concentration difference is needed to keep diffusion happening rapidly. Ventilation continuously replaces alveolar air, while circulation continuously removes oxygenated blood. Together, these maintain the gradient between the two sides of the surface.

MisconceptionMany students confuse ‘large surface area’ with ‘large surface area to volume ratio’. A whale has a huge surface area but a small SA:V ratio. The ratio matters for diffusion, not absolute size.
Exam TipAlways specify SA:V ratio, not just surface area.
Fick’s Law of Diffusion
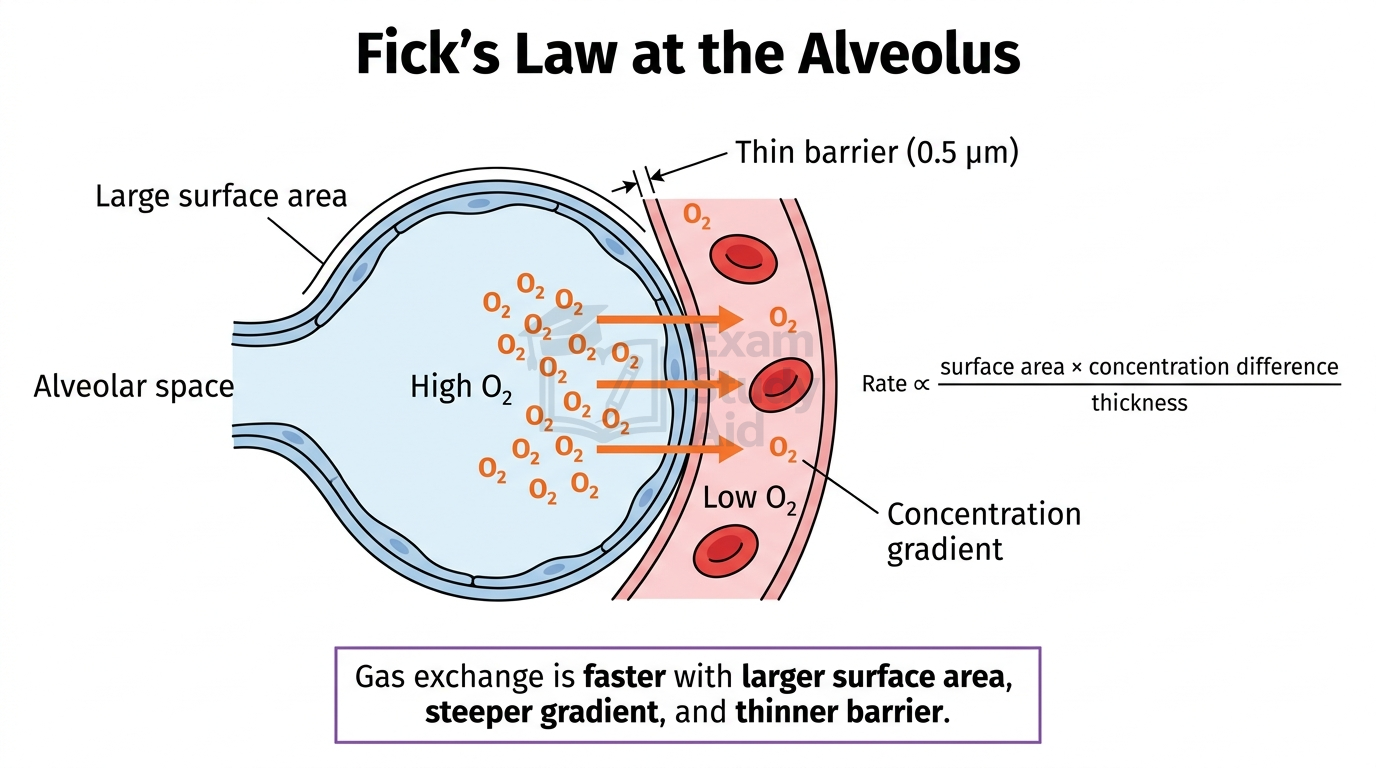
Fick’s Law of Diffusion quantifies how the rate of diffusion across an exchange surface depends on its three key properties.
The law is expressed as:
Rate of diffusion ∝ (surface area × concentration difference) ÷ thickness of membrane
The three variables work as follows:
- Surface area: a larger surface allows more molecules to diffuse simultaneously.
- Concentration difference: a steeper gradient drives faster net movement.
- Thickness: a thinner barrier shortens the diffusion distance.
Each property has a predictable effect. Doubling the surface area doubles the rate of diffusion. Doubling the concentration difference also doubles the rate. Halving the thickness doubles the rate.
For example, if alveolar walls thicken from 0.5 µm to 1.0 µm due to fibrosis, the diffusion rate halves. This assumes surface area and concentration difference remain constant. This is why diseases that damage exchange surfaces seriously impair gas exchange.
Examiner InsightWhen asked how a structure increases the rate of diffusion, link directly to Fick’s Law. State which variable changes (surface area, concentration difference, or thickness) and how that change increases the rate.
Exam TipAlways state the Fick’s Law variable explicitly.
Mammalian Lung Structure
The mammalian lung shows multiple structural adaptations that maximise the rate of gas exchange between alveolar air and the blood capillary network.
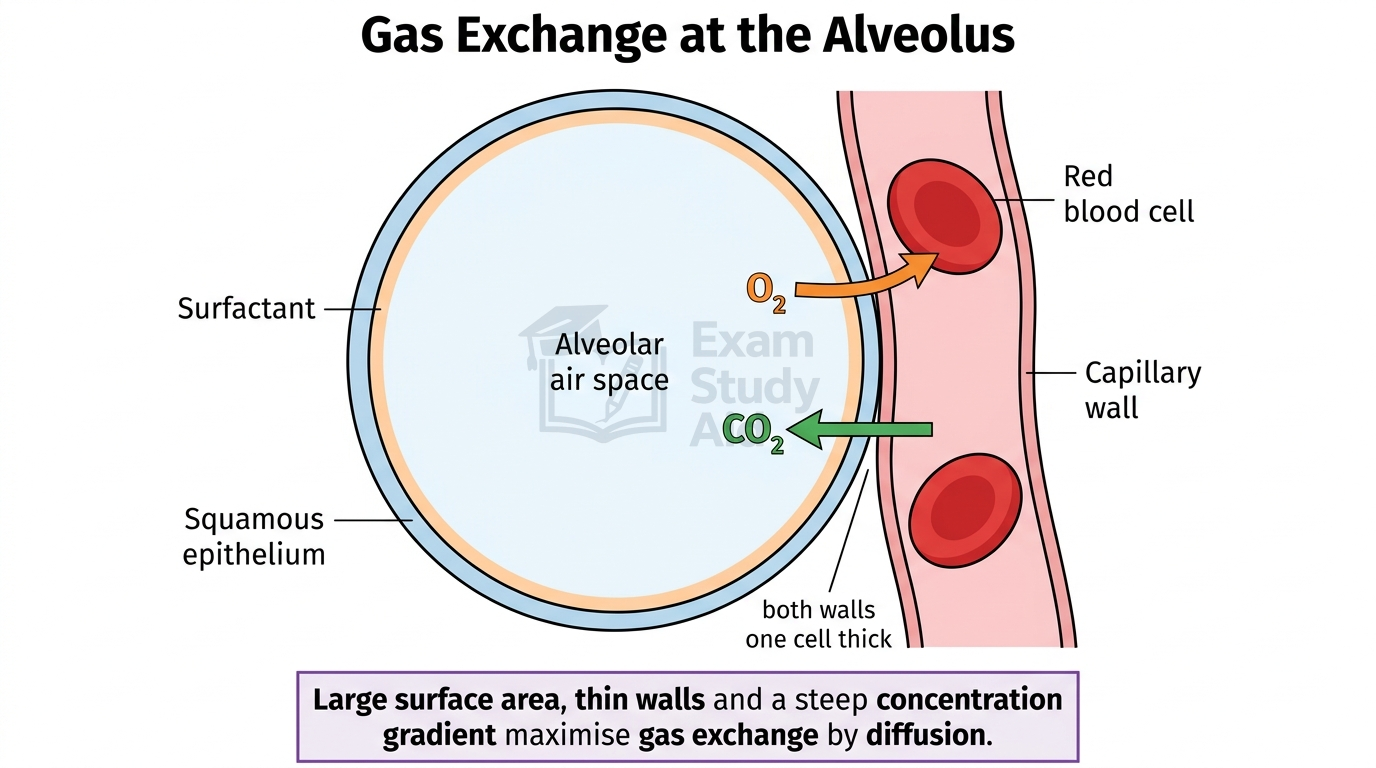
The lungs contain millions of tiny air sacs called alveoli, each surrounded by a dense network of capillaries. Together, these provide an enormous surface area of around 70 m² in an adult human, while keeping the diffusion distance very short.
| Adaptation | How it increases gas exchange efficiency |
|---|---|
| Millions of alveoli | Provide a very large total surface area (~70 m²) for diffusion |
| Alveolar wall one cell thick | Made of squamous epithelium, reducing diffusion distance |
| Capillary wall one cell thick | Made of endothelium, also reducing diffusion distance |
| Dense capillary network | Brings blood close to every alveolus |
| Continuous ventilation | Replaces alveolar air, maintaining a steep concentration gradient |
| Continuous blood flow | Carries oxygenated blood away and brings deoxygenated blood in |
| Surfactant lining | Reduces surface tension, prevents alveolar collapse |
| Moist alveolar surface | Allows gases to dissolve before diffusing into blood |
Each of these adaptations maps directly onto Fick’s Law. Alveoli increase surface area, the thin walls decrease thickness, and ventilation plus blood flow maintain the concentration difference. Together, they make the lung extraordinarily efficient at supplying the body with oxygen and removing carbon dioxide.
Cell Membrane Structure and Properties
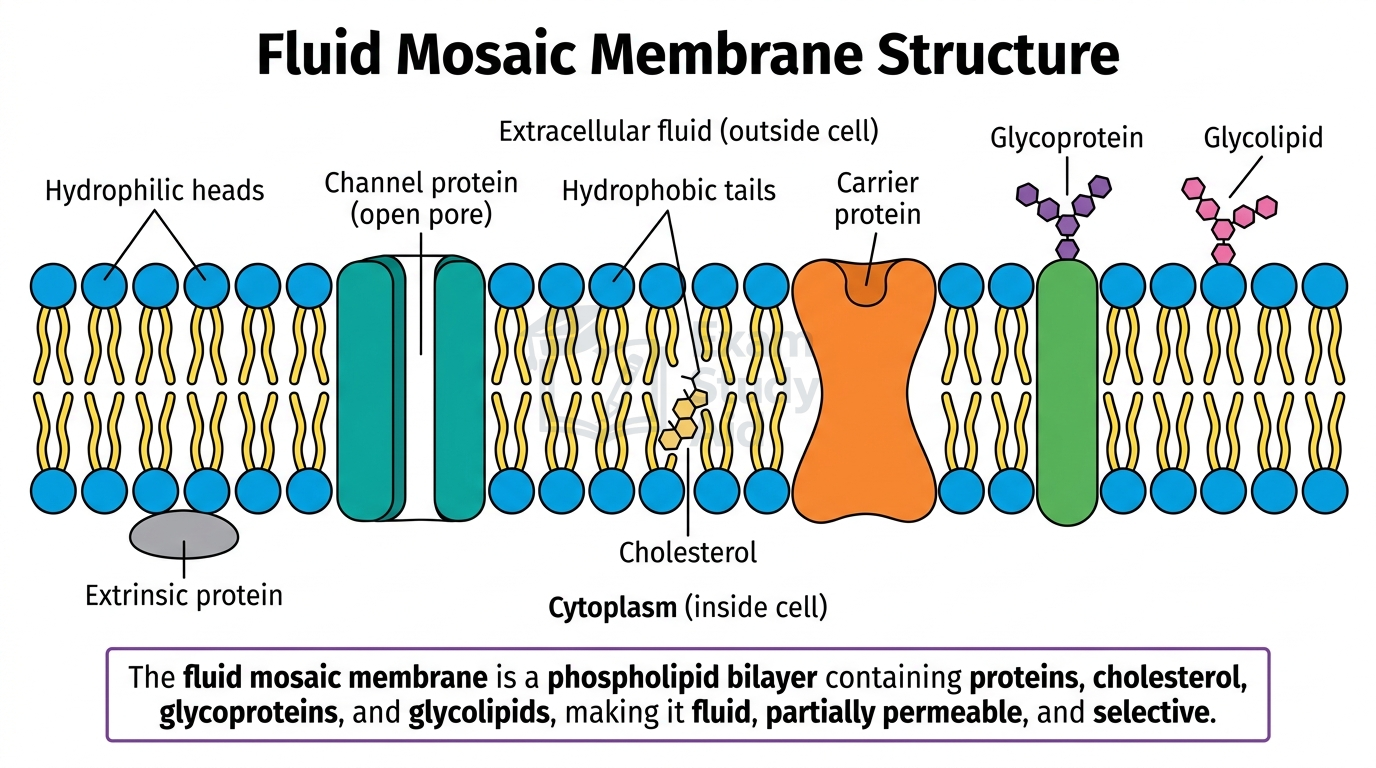
Cell membranes are partially permeable barriers built from a phospholipid bilayer with proteins, cholesterol, and carbohydrates embedded throughout the structure.
Each phospholipid has a hydrophilic [water-attracting] phosphate head and two hydrophobic [water-repelling] fatty acid tails. They arrange themselves spontaneously in water, with heads facing outward toward the aqueous environment and tails pointing inward, away from water. This bilayer forms a barrier to water-soluble molecules and ions.
The structure of the membrane includes several key components, each with a specific role:
| Component | Description | Role |
|---|---|---|
| Phospholipid bilayer | Hydrophilic heads outward; hydrophobic tails inward | Forms the basic structure; barrier to water-soluble molecules and ions |
| Intrinsic (integral) proteins | Span the entire bilayer | Act as channel or carrier proteins; some function as enzymes or receptors |
| Extrinsic (peripheral) proteins | Attached to one surface of the bilayer | Act as receptors or provide structural support |
| Cholesterol | Wedged between phospholipid molecules | Regulates fluidity, stabilising the membrane across temperatures |
| Glycoproteins | Proteins with carbohydrate chains attached, on the outer surface | Involved in cell recognition and cell signalling |
| Glycolipids | Lipids with carbohydrate chains attached, on the outer surface | Involved in cell recognition and cell signalling |
The membrane has three key properties. It is fluid because phospholipids can move laterally within the bilayer. It is partially permeable because only certain molecules cross freely without help. It is selective because specific transport proteins control which molecules enter or leave the cell.
The Fluid Mosaic Model
The fluid mosaic model, proposed by Singer and Nicolson in 1972, describes the membrane as a fluid lipid bilayer with proteins distributed throughout like tiles in a mosaic.
The model is called fluid because the phospholipid molecules can move laterally within the bilayer, much like molecules in a liquid layer. It is called mosaic because the proteins are scattered throughout the bilayer in an irregular pattern.
This model results from decades of evidence and has replaced older models like the Davson-Danielli sandwich model. That older model proposed proteins formed continuous layers on either side of the bilayer, like bread in a sandwich. New evidence overturned that model, showing models in science are interpretations of data, refined as new observations emerge.
Key evidence for the fluid mosaic model includes:
- Freeze-fracture electron microscopy splits the bilayer and reveals proteins embedded throughout, not as continuous layers.
- Fluorescent labelling experiments: when mouse and human cell membranes were fused, their labelled proteins mixed within minutes, proving membrane proteins move laterally.
- Differential staining showed proteins penetrate into the bilayer, not just sitting on its surface.
This illustrates how scientific models develop. Initial proposals are tested against new techniques and data. Models that fail to explain new evidence are revised or replaced. The fluid mosaic model continues to be supported by current data and remains the accepted explanation.
Examiner InsightExaminers testing the fluid mosaic model expect both descriptive accuracy and supporting evidence. State that the membrane is fluid because phospholipids move laterally, and mosaic because proteins are scattered. Then link to evidence such as freeze-fracture or fluorescent labelling.
Exam TipUse the words ‘fluid’ and ‘mosaic’ explicitly, with their meanings.
Investigating Membrane Permeability
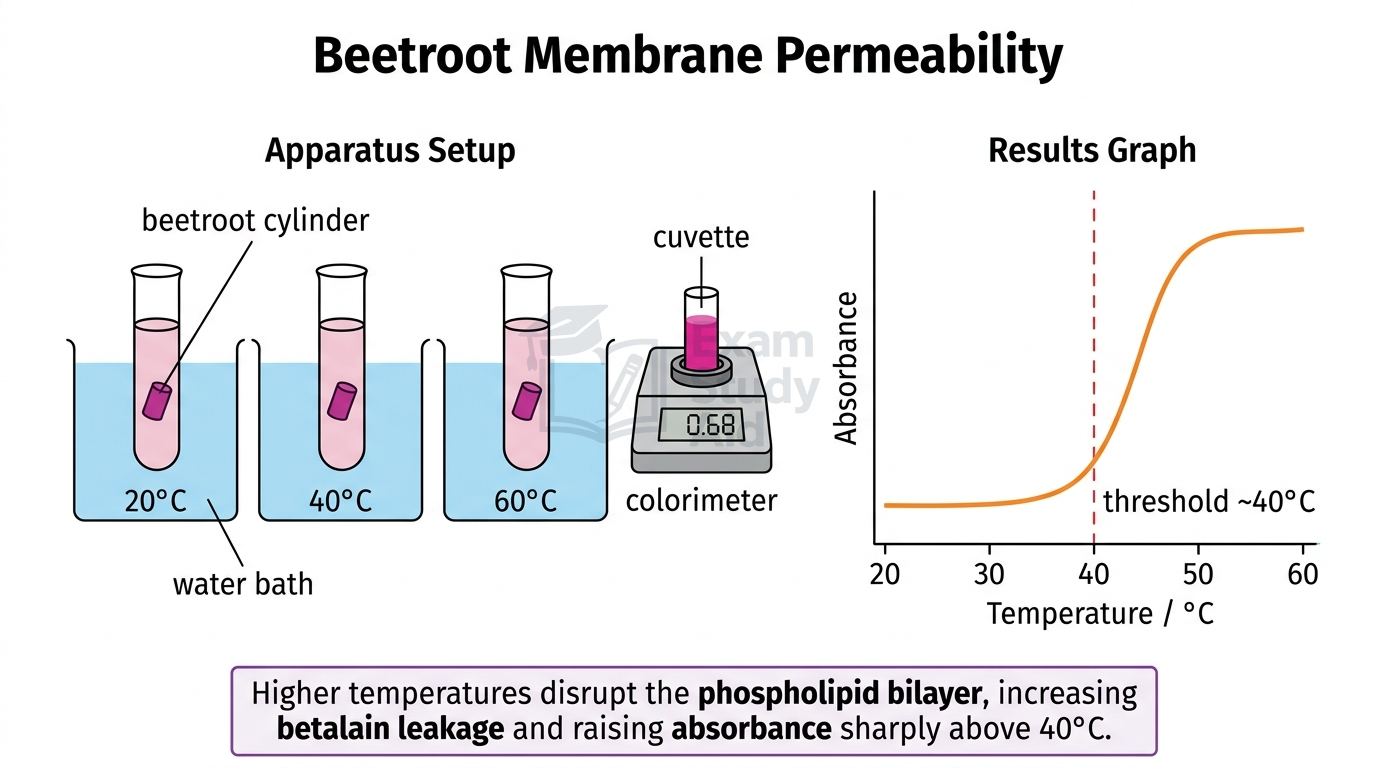
Membrane permeability can be measured by tracking how much pigment leaks out of beetroot cells when conditions disrupt the phospholipid bilayer.
Beetroot vacuoles contain a red pigment called betalain [a water-soluble red pigment]. The cell surface membrane and tonoplast normally trap this pigment inside. If these membranes are damaged, betalain leaks into the surrounding solution. The amount of leakage is measured using a colorimeter [an instrument measuring light absorbance through a sample], with higher absorbance meaning more pigment release and therefore greater membrane permeability.
CORE PRACTICAL 3: Effect of alcohol and temperature on membrane permeability
Aim: To investigate how alcohol concentration and temperature affect cell surface membrane permeability using beetroot.
- Use a cork borer to cut beetroot cylinders of identical diameter, then trim each to the same length (e.g. 1 cm).
- Wash the cylinders thoroughly under running water to remove pigment from cut surfaces.
- For temperature investigation: place one cylinder per test tube containing 5 cm³ distilled water. Place tubes in water baths at 10°C, 20°C, 30°C, 40°C, 50°C, and 60°C for 30 minutes.
- For alcohol investigation: place one cylinder per test tube containing 5 cm³ of ethanol. Use concentrations of 0%, 20%, 40%, 60%, and 80%, then leave at room temperature for 30 minutes.
- Remove the beetroot, agitate the solution, and transfer to a cuvette.
- Use a colorimeter (with a green/blue filter, ~530 nm) to measure absorbance of each solution.
- Repeat at least three times per condition and calculate the mean.
Independent variable (IV): Either temperature in °C or alcohol concentration in %. Dependent variable (DV): Absorbance of the solution at 530 nm (proportional to pigment leakage). Control variables (CV): Volume of water/alcohol, size of beetroot cylinder, time submerged, beetroot variety, source of beetroot.
As temperature increases, absorbance stays low up to ~40°C, then rises sharply because heat denatures membrane proteins and increases phospholipid fluidity, creating gaps. As alcohol concentration increases, absorbance rises because alcohol dissolves the phospholipid bilayer, disrupting its structure.
All beetroot cylinders must be the same size, since differences in surface area would change leakage rates and confound the IV. Wear gloves to avoid staining hands and a lab coat to protect clothing.
QUICK RECAP
Key Points
- Gas exchange surfaces share three properties: large SA:V, thin barrier, steep concentration gradient.
- Fick’s Law: rate of diffusion ∝ (surface area × concentration difference) ÷ thickness.
- Doubling surface area or halving thickness doubles the rate of diffusion.
- Mammalian lung has ~70 m² of alveolar surface area.
- Alveolar wall is one squamous epithelium cell thick.
- Capillary wall is one endothelium cell thick.
- Ventilation and blood flow maintain steep concentration gradients.
- Surfactant reduces surface tension and prevents alveolar collapse.
- Cell membranes are partially permeable barriers with a phospholipid bilayer.
- Hydrophilic phosphate heads face outward; hydrophobic fatty acid tails face inward.
- Intrinsic proteins span the bilayer; extrinsic proteins attach to one side.
- Cholesterol regulates membrane fluidity across temperature ranges.
- Glycoproteins and glycolipids enable cell signalling and recognition.
- Fluid mosaic model: phospholipids move laterally, proteins scattered throughout.
- Evidence includes freeze-fracture microscopy and fluorescent labelling experiments.
- Beetroot pigment leaks when temperature or alcohol disrupts the membrane.
- Colorimeter measures absorbance at 530 nm to quantify pigment release.
- High temperatures denature membrane proteins and increase fluidity.
- Alcohol dissolves the phospholipid bilayer, increasing permeability.
CAN I…? PROGRESS CHECK
Self-Assessment
- …state the three properties of an efficient gas exchange surface?
- …write Fick’s Law and apply it to a calculation question?
- …describe at least four adaptations of the mammalian lung for gas exchange?
- …label all the components of the fluid mosaic membrane structure?
- …explain the role of cholesterol, glycoproteins, and glycolipids?
- …describe two pieces of evidence supporting the fluid mosaic model?
- …identify variables and controls in the beetroot permeability investigation?
- …explain why high temperature and alcohol increase membrane permeability?